Early, often overlooked psychological conjecture emphasizes that spontaneous behavioral variability is a useful, as one would say today “adaptive” trait. In this article I will cite neurobiological evidence to strengthen this view. I will use a number of examples to argue that the variability measured in the behavioral performance of animals is exactly the kind of output that is required to effectively detect which of the stimuli in the incoming stream of sensory input can be controlled by the animal and which cannot. I will deliver an account as to how and why, despite its importance, this essential output-input feature of brains has largely been overlooked in recent decades. This forgotten feature is associated with a number of psychiatric disorders and only recently a new and growing trend has emerged which now provides steadily increasing understanding about the mechanisms underlying it.
Behavioral Variability: the output
We all feel the very basic notion that we possess a certain degree of freedom of choice. Bereaving humans of such freedom is frequently used as punishment and the bereft do in-variably perceive this limited freedom as undesirable. This experience of freedom is an important characteristic of what it is like to be human. It stems in part from our ability to behave variably. Voltaire expressed this intuition in saying “Liberty then is only and can be only the power to do what one will” [2]. But the concept that we can decide to behave differently even under identical circumstances underlies not only our justice systems. Electoral systems, our educational systems, parenting and basically all other social systems also presuppose behavioral variability and at least a certain degree of freedom of choice. Games and sports would be predictable and boring without our ability of constantly changing our behavior in always the same settings. Faced with novel situations, humans and most animals spontaneously increase their behavioral variability [3-5]. Inasmuch as behavioral variability between individuals has genetic components, it is a crucial factor of niche exploitation in evolution. Moreover, behavioral variability within individuals has been shown to be ecologically advantageous in game theoretical studies [6-11], in pursuit-evasion contests such as predator/prey interactions (“Protean Strategy”) [12-15], in exploration/foraging [16], in mobbing attack patterns by birds and in the variation of male songbirds’ songs [17]. Clearly, invariable behavior will be exploited [14,18] and leaves an organism helpless in unpredictable situations [19,20].
Controlling external events: the input
Thus, competitive success and evolutionary fitness of all ambulatory organisms rely critically on intact behavioral variability as an adaptive brain function. But relative freedom from environmental contingencies is a necessary, but most often not a sufficient criterion for such accomplishments. Tightly connected to the ability to produce variable behavior is the ability to use the effects of these behaviors to control the environment. The incoming stream of sensory information is noisy and fluctuates for any number of reasons. Any covariance between the behavioral variations and those of sensory input indicates that the latter are con-sequences of the behavior and can thus be controlled be the animal [21,22]. It is the on-line detection system for when the animal itself is the reason for any environmental fluctuation. This function is so paramount, that we humans express our delight over control of our environment (including other people) already as children, by e.g., shrieking in excitement when Daddy jumps after a “boo” or proudly presenting Mom with “look what I can do!”. Later, children find pleasure in building airplane models, become carpenters with a delight for shaping wood, artists feeling gratified creating art out of the simplest materials, musicians enjoying mastering their instrument to perfection, athletes, scientists, engineers or managers. Using trial and error, we have shaped our world from caves to skyscrapers, from horses to jet-planes, from spears to hydrogen bombs. Cultural or religious rituals (e.g., rain dance) and superstition may have evolved as means to create a feeling of control where ultimately there is none. Obviously, behaving flexibly in order to control our environment is at the heart of human nature and probably affects more aspects of our daily lives than any other cognitive brain function. So essential is such functioning that even very simple brains possess it. The modest fruit fly prefers a situation in which it controls its environment over one where it does not. If certain flight directions are experimentally superimposed with uncontrollable visual movements, flies quickly avoid such directions and fly only in areas of full control [23]. This experiment demonstrates that control over environmental stimuli is inherently rewarding already for simple, but more likely for all brains.
The main function of brains
The first experiments into the mechanistic basis of this basic brain function was initiated already early in the 20th century by eminent scientists like Thorndike [24], Watson [25] and Skinner [26]. Of course, the primary process by which all animals, including humans learn to control their environment is operant (or instrumental) conditioning (Box 1). Ultimately, this comparatively simple process forms one of the fundamental cornerstones not only for all of our human nature, but also for our social coherence: human nature as described in planning, willing and controlling our behavior [22,27-30] and our social coherence as based on cooperation [6,31,32]. Modern neuroscience, however, with the success of research into the mechanisms of the even simpler process of Pavlovian or classical conditioning (Box 1), has understandably shifted the focus away from the central role operant learning plays in our daily lives.
| Box 1: Predictive learning Classical (Pavlovian) conditioning is the process by which we learn the relationship between events in our environment, e.g., that lightning always precedes thunder. The most famous classical conditioning experiment involves Pavlov’s dog: The physiologist I.P. Pavlov trained dogs to salivate in anticipation of food by repeatedly ringing a bell (conditioned stimulus, CS) before giving the animals food (unconditioned stimulus, US). Dogs naturally salivate to food. After a number of such presentations, the animals would salivate to the tone alone, indicating that they were expecting the food. Operant (instrumental) conditioning is the process by which we learn about the consequences of our actions, e.g. not to touch a hot plate. The most famous operant conditioning experiment involves the ‘Skinner-Box’ in which the psychologist B.F. Skinner (and colleagues – he mainly used pigeons himself) trained rats to press a lever for a food reward. The animals were placed in the box and after some exploring would also press the lever, which would lead to food pellets being dispensed into the box. The animals quickly learned that they could control food delivery by pressing the lever. Both operant and classical conditioning serve to be able to predict the occurrence of important events (such as food). However, one of a number of important differences in particular suggests that completely different brain functions underlie the two processes. In classical conditioning external stimuli control the behavior by triggering certain responses. In operant conditioning the behavior controls the external events.
|

This shift is signified by a steady decrease in the fraction of biomedical publications with operant topics, despite an absolute increase of publications over the last 25 years (Fig. 1). It is an understandable shift, because nearly every learning situation seems to involve a dominant classical component anyway [33,34] and classical conditioning offers the unique advantage to quickly and easily get at the biological processes underlying learning and memory: the animals are usually restrained, leaving only few degrees of freedom and the stimuli can be traced to the points of convergence where the learning has to take place. The neurobiological study of classical conditioning, pioneered by Nobel laureate Eric Kandel, was the first avenue into some of the biological mechanisms of general brain function. Today, overwhelmed by the amazing progress in the past three decades, some neuroscientists even ponder reducing general brain function almost exclusively (“95%”) to classical stimulus-response relationships, with profound implications for society, in particular for the law [35,36]. The Dana Foundation, the American Association for the Advancement of Science and the American Civil Liberties Union have already sponsored meetings on these implications [37,38]. Stretching the generality of such awesome classical conditioning paradigms as fear conditioning in rats and mice [39], rabbit eyeblink conditioning [40] or classical conditioning of the Aplysia gill withdrawal reflex [41], the current neuroscientific standard implies that they are all-encompassing paradigms for general cognitive brain function: “brain function is ultimately best understood in terms of input/output transformations and how they are produced” [42].
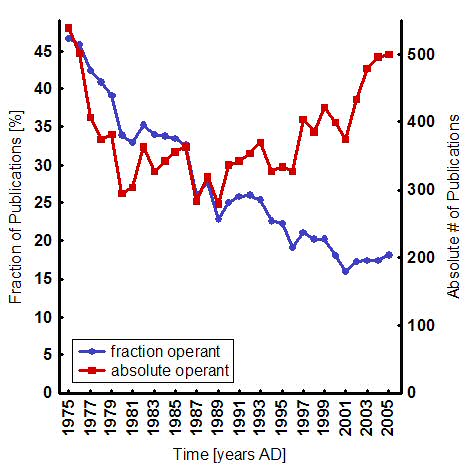 Fig.1: Publishing development of publications on operant conditioning. The graph shows a steady decline in the fraction of publications dealing with operant conditioning (blue) despite an increase in absolute number of publications over the last 25 years (red). Note the increase in the last four years. Absolute numbers were gathered by running an NCBI PubMed query “((operant OR instrumental) AND (conditioning OR learning))” (red). This number was divided by the number of publications containing only “conditioning”, to derive a percentage (blue). Notice the sharp jump in the absolute number for the last five years. This jump is even noticeable in the relative contributions. |
It is rarely recognized that, at an adaptive level, cognitive capacities, such as those involved in encoding the predictive relations between stimuli, can be of little functional value to a hypothetical, purely Pavlovian organism. For instance, one can imagine any number of situations which require the animal to modify, even to withhold or reverse, the direction of some behavior in order to solve the situation. Such situations demand greater behavioral flexibility than the system mediating classical conditioning provides. Moreover, using the re-afference principle [43-45], operant behavior underlies the distinction between observing and doing, i.e. differentiating between self and non-self. We compare our behavioral output (efference) with incoming sensory input (afference) to detect when we are the ones authoring environmental change [21,22]. One almost iconographic example of such behavior is to per-form various spontaneous movements in front of a mirror to detect whether it is us we are perceiving [46,47]. This automatic detection-mechanism explains why we cannot tickle our-selves [21], why we perceive a stable visual world despite our frequent quick, or saccadic, eye movements [48] and is reflected in different brain activation patterns between self-generated and exogenous visual stimulation [49]. It is thought that the detection is accomplished via an efference copy (or corollary discharge) of the motor command which is compared to incoming afferent signals to distinguish re-afference from ex-afference. Such a differentiation has been implied to demonstrate causal reasoning in rats [50,51]. Even robots can use such “self-modeling” to generate a continuously updated model of themselves and their environment [52]. Conspicuously, the organization of the brain also raises doubts about the input/output mainstream image. Less than 10% of all synapses in the brain carry incoming sensory information and as little as 0.5-1% of the brain’s total energy budget are sufficient to handle the momentary demands of the environment [53]. In other words, input/output transformations may only account for a small fraction of what brains are doing. Maybe a much more significant portion of the brain is occupied with the ongoing modeling of the world and how it might react to our actions? Recent evidence suggests that the brain predicts the sensory consequences of motor commands because it integrates its prediction with the actual sensory information to produce an estimate of sensory space that is enhanced over predictions from either source alone [54]. This effect of operant enhancement of sensory cues can be observed also in fruit fly learning [23,34] and may explain why starlings, but not tamarin monkeys can recognize patterns defined by so-called recursive grammar [55]. Such control of sensory input has often been termed “goal-directed” behavior. This perspective provides an intuitive under-standing of the rewarding properties of being in control of the environment. Setting and obtaining goals is inherently rewarding [56]. This reward ensures that individuals always actively strive to control.
Far-reaching implications
This evidence indicates that our behavior consists at least as much of goal-directed actions as it consists of responses elicited by external stimuli. But not all stimulus-response contingencies are acquired by classical conditioning. Goal-directed actions can become partially independent of their environmental feedback and develop into habits controlled mainly by antecedent stimuli [62-64]. Everybody has experienced such ‘slip of action’ instances, when we take the wrong bus home days after we have moved, when we keep reaching for the wrong buttons or levers in our new car, when we try to open our home door with the work keys or when we take the freeway-exit to our workplace, even though we were heading for the family retreat. William James [65] is often quoted as claiming that “very absent-minded persons in going in their bedroom to dress for dinner have been known to take of one garment after an-other and finally to get into bed, merely because that was the habitual issue of the first few movements when performed at a late hour”.
The experience of willing to do something and then successfully doing it is absolutely central to developing a sense of who we are (and who we are not) and that we are in control (and not being controlled). This sense is compromised in patients with dissociative identity disorder, alien hand syndrome, or schizophrenic delusions [21]. In some of these disorders the abovementioned midbrain dopamine neurons appear to play a central role, tying, e.g., Parkinson and schizophrenia tightly to operant models. Parkinson’s patients are administered the dopamine precursor L-DOPA, while schizophrenics are treated with a group of antipsychotics, most of which target and inhibit the D2 dopamine receptor. Some of these antipsychotic drugs have Parkinson-like side-effects. Recent research shows that L-DOPA and the antipsychotic haloperidol have opposite effects on operant decision-making in humans [83]. One is tempted to interpret these data as evidence for the hypothesis that the overlapping and interacting dopaminergic systems mediating primary rewards such as food, water or sex and those mediating behavior initiation and control are so tightly inter-connected precisely because of the rewarding properties of controlling the environment with behavior. As information such as the above accumulates, elucidating the mechanisms of operant conditioning becomes more and more promising as an avenue into understanding the causality underlying disorders such as those described above and their treatment.
Scarce but converging biological data
Compared to its significance, our understanding of the biological mechanisms underlying operant conditioning is rather vague. The more important is a recent swell of ground-breaking studies (see also Fig. 1). A number of different model systems have contributed to this progress on various levels of operant conditioning. I will try to integrate the knowledge gained from such disparate sources to describe the general picture as it is currently emerging.
Another case for multiple model systems
Our relative lack of knowledge stems in part from research into operant conditioning being conceptually much more challenging than classical conditioning. However, recent progress in invertebrate neuroscience suggests that the now classic Kandelian approach of relying heavily on simpler brains while developing tools and models for vertebrate research is even more promising today in the age of advanced molecular, genetic, imaging and physiological repertoires in invertebrates than 30 years ago [20,106]. Even in the post-genomic era, invertebrate models offer the possibility to rapidly and effectively learn about important principles and molecules which can then be used to reduce the complexity of the vast vertebrate brain [107]. Besides offering a more effective avenue into studying the neural basis of operant conditioning, such an integrative approach will provide us with insights into the exciting question of why invertebrate and vertebrate brains are structurally so very different even though the basic demands of life are quite similar in both groups. Moreover, a multi-faceted approach will allow us to distinguish general mechanisms from species-specific adaptations. Coincidentally, using multiple model systems effectively reduces the number of vertebrate experimental animals, working towards the ‘3R’ goals — refinement, reduction and replacement [108]. Combining the rapid technical advancements also in vertebrate physiology, imaging and behavior [109] with modern computational power, neuroscience is now more than ready to finally tackle operant conditioning on a broad scale. The recent swell of publications on operant conditioning is the logical consequence of 20 years of meticulous research during the dominant input/output mainstream [62]. The most important questions to be answered in future research are:
- What are the brain-circuits generating spontaneous behavior?
- How is sensory feedback integrated into these circuits?
- Which are the ‘operant’ genes?
- What are the mechanisms by which fact-learning suppresses skill-learning?
- How can repetition overcome this suppression?
Acknowledgments: I am grateful to Bernd Grünewald, Bernhard Komischke, Gérard Leboulle, Diana Pauly, John Caulfield, Peter Wolbert, Martin Heisenberg and Randolf Menzel for critically reading an earlier version of the manuscript. I am especially indebted to Bernard Balleine and Charles Beck for providing encouragement, stimulating information and some key references.
References:
1. Kant I (1781) Critique of Pure Reason: NuVision Publications, LLC. 444 p.
2. Voltaire (1752/1924) Voltaire’s philosophical dictionary. Wolf HI, translator. New York, NY, USA: Knopf.
3. Shahan TA, Chase PN (2002) Novelty, stimulus control, and operant variability. Behavior Analyst 25: 175-190.
4. Roberts S, Gharib A (2006) Variation of bar-press duration: Where do new responses come from? Behavioural Processes
Proceedings of the Meeting of the Society for the Quantitative Analyses of Behavior – SQAB 2005 72: 215-223.
5. Bunzeck N, Duzel E (2006) Absolute Coding of Stimulus Novelty in the Human Substantia Nigra/VTA. Neuron 51: 369-379.
6. McNamara JM, Barta Z, Houston AI (2004) Variation in behaviour promotes cooperation in the Prisoner’s Dilemma game. Nature 428: 745-748.
7. Glimcher P (2003) Decisions, uncertainty, and the brain: the science of neuroeconomics. Cambridge, MA: MIT.
8. Glimcher PW, Rustichini A (2004) Neuroeconomics: the consilience of brain and decision. Science 306: 447-452.
9. Glimcher PW (2005) Indeterminacy in brain and behavior. Annu Rev Psychol 56: 25-56.
10. Platt ML (2004) Unpredictable primates and prefrontal cortex. Nat Neurosci 7: 319-320.
11. Brembs B (1996) Chaos, cheating and cooperation: Potential solutions to the Prisoner’s Dilemma. Oikos 76: 14-24.
12. Grobstein P (1994) Variability in behavior and the nervous system. In: Ramachandran VS, editor. Encyclopedia of Human Behavior. New York: Academic Press. pp. 447-458.
13. Driver PM, Humphries N (1988) Protean behavior: The biology of unpredictability. Ox-ford, England: Oxford University Press.
14. Miller GF (1997) Protean primates: The evolution of adaptive unpredictability in competition and courtship. In: Whiten A, Byrne RW, editors. Machiavellian Intelligence II: Extensions and evaluations. Cambridge: Cambridge University Press. pp. 312-340.
15. Shultz S, Dunbar R (2006) Chimpanzee and felid diet composition is influenced by prey brain size. Biology Letters: FirstCite.
16. Belanger JH, Willis MA (1996) Adaptive control of odor-guided locomotion: Behavioral flexibility as an antidote to environmental unpredictability. Adaptive Behavior 4: 217-253.
17. Neuringer A (2004) Reinforced variability in animals and people: implications for adaptive action. Am Psychol 59: 891-906.
18. Jablonski PG, Strausfeld NJ (2001) Exploitation of an ancient escape circuit by an avian predator: relationships between taxon-specific prey escape circuits and the sensitivity to visual cues from the predator. Brain Behav Evol 58: 218-240.
19. Heisenberg M (1994) Voluntariness (Willkürfähigkeit) and the general organization of behavior. Life Sciences Research Report 55: 147-156.
20. Greenspan RJ (2005) No Critter Left Behind: An Invertebrate Renaissance. Current Biology 15: R671-R672.
21. Bays PM, Flanagan JR, Wolpert DM (2006) Attenuation of Self-Generated Tactile Sensations Is Predictive, not Postdictive. PLoS Biology 4: e28.
22. Wegner DM (2002) The illusion of conscious will. Boston: Bradford Books/MIT press. 419 p.
23. Heisenberg M, Wolf R, Brembs B (2001) Flexibility in a single behavioral variable of Drosophila. Learn Mem 8: 1-10.
24. Thorndike EL (1911) Animal Intelligence. New York: Macmillan.
25. Watson J (1928) The ways of behaviorism. New York: Harper & Brothers Pub.
26. Skinner BF (1938) The behavior of organisms. New York: Appleton.
27. Owen AM (1997) Cognitive planning in humans: neuropsychological, neuroanatomical and neuropharmacological perspectives. Prog Neurobiol 53: 431-450.
28. Frith CD, Friston K, Liddle PF, Frackowiak RS (1991) Willed action and the prefrontal cortex in man: a study with PET. Proc R Soc Lond B Biol Sci 244: 241-246.
29. Knight RT, Grabowecky MF, Scabini D (1995) Role of human prefrontal cortex in attention control. Adv Neurol 66: 21-34; discussion 34-26.
30. Lezak MD (1995) Neuropsychological Assessment. New York, NY: Oxford University Press. 1056 p.
31. Gutnisky DA, Zanutto BS (2004) Cooperation in the iterated prisoner’s dilemma is learned by operant conditioning mechanisms. Artificial Life 10: 433-461.
32. Sanabria F, Baker F, Rachlin H (2003) Learning by pigeons playing against tit-for-tat in an operant prisoner’s dilemma. Learn Behav 31: 318-331.
33. Rescorla RA (1987) A Pavlovian analysis of goal-directed behavior. American Psychologist 42: 119-129.
34. Brembs B (2006, subm.) Hierarchical interactions facilitate Drosophila predictive learning. Neuron.
35. Greene J, Cohen J (2004) For the law, neuroscience changes nothing and everything. Philos Trans R Soc Lond B Biol Sci 359: 1775-1785.
36. Weigmann K (2005) Who we are. EMBO Rep 6: 911-913.
37. Garland B, Glimcher PW (2006) Cognitive neuroscience and the law. Current Opinion in Neurobiology 16: 130-134.
38. Garland B (2004) Neuroscience and the Law: Brain, Mind, and the Scales of Justice. Chicago: University of Chicago Press.
39. LeDoux JE (2000) Emotion Circuits in the Brain. Annual Review of Neuroscience 23: 155-184.
40. Medina JF, Christopher Repa J, Mauk MD, LeDoux JE (2002) Parallels between cerebellum- and amygdala-dependent conditioning. Nat Rev Neurosci 3: 122-131.
41. Kandel ER (2001) The molecular biology of memory storage: a dialogue between genes and synapses. Science 294: 1030-1038.
42. Mauk MD (2000) The potential effectiveness of simulations versus phenomenological models. Nat Neurosci 3: 649-651.
43. Todorov E (2004) Optimality principles in sensorimotor control. Nat Neurosci 7: 907-915.
44. von Holst E, Mittelstaedt H (1950) Das Reafferenzprinzip. Wechselwirkungen zwischen Zentralnervensystem und Peripherie. Naturwissenschaften: 464-476.
45. Webb B (2004) Neural mechanisms for prediction: do insects have forward models? Trends Neurosci 27: 278-282.
46. Plotnik JM, de Waal FBM, Reiss D (2006) Self-recognition in an Asian elephant. PNAS 103: 17053-17057.
47. Reiss D, Marino L (2001) Mirror self-recognition in the bottlenose dolphin: A case of cognitive convergence. PNAS 98: 5937-5942.
48. Sommer MA, Wurtz RH (2006) Influence of the thalamus on spatial visual processing in frontal cortex. Nature advanced online publication.
49. Matsuzawa M, Matsuo K, Sugio T, Kato C, Nakai T (2005) Temporal relationship be-tween action and visual outcome modulates brain activation: an fMRI study. Magn Reson Med Sci 4: 115-121.
50. Blaisdell AP, Sawa K, Leising KJ, Waldmann MR (2006) Causal Reasoning in Rats. Science 311: 1020-1022.
51. Clayton N, Dickinson A (2006) Rational rats. Nat Neurosci 9: 472-474.
52. Bongard J, Zykov V, Lipson H (2006) Resilient Machines Through Continuous Self-Modeling. Science 314: 1118-1121.
53. Raichle ME (2006) NEUROSCIENCE: The Brain’s Dark Energy. Science 314: 1249-1250.
54. Vaziri S, Diedrichsen J, Shadmehr R (2006) Why Does the Brain Predict Sensory Consequences of Oculomotor Commands? Optimal Integration of the Predicted and the Ac-tual Sensory Feedback. J Neurosci 26: 4188-4197.
55. Marcus GF (2006) Language: Startling starlings. Nature 440: 1117-1118.
56. Kim H, Shimojo S, O’Doherty JP (2006) Is Avoiding an Aversive Outcome Rewarding? Neural Substrates of Avoidance Learning in the Human Brain. PLoS Biology 4: e233.
57. Duckworth R (2006) Aggressive behaviour affects selection on morphology by influencing settlement patterns in a passerine bird. Proc R Soc Lond B: FirstCite.
58. Dickens WT, Flynn JR (2001) Heritability estimates versus large environmental effects: the IQ paradox resolved. Psychol Rev 108: 346-369.
59. Toga AW, Thompson PM (2005) Genetics of brain structure and intelligence. Annu Rev Neurosci 28: 1-23.
60. Gandevia SC, Smith JL, Crawford M, Proske U, Taylor JL (2006) Motor commands contribute to human position sense. J Physiol (Lond) 571: 703-710.
61. Gintis H (2006, in press) A Framework for the Unification of the Behavioral Sciences. Behavioral and Brain Sciences.
62. Yin HH, Knowlton BJ (2006) The role of the basal ganglia in habit formation. Nat Rev Neurosci 7: 464-476.
63. Hernandez PJ, Schiltz CA, Kelley AE (2006) Dynamic shifts in corticostriatal expression patterns of the immediate early genes Homer 1a and Zif268 during early and late phases of instrumental training. Learn Mem 13: 599-608.
64. Aragona BJ, Carelli RM (2006) Dynamic neuroplasticity and the automation of motivated behavior. Learn Mem 13: 558-559.
65. James W (1890) The Principles of Psychology. New York: Holt.
66. Davis H. fMRI Bold Signal changes in Athletes in response to video review of failure: Effects of cognitive reappraisal (CBT). 2006; San Francisco.
67. Seligman M (1975) Helplessness: On depression, development, and death.: W. H. Free-man.
68. Amat J, Baratta MV, Paul E, Bland ST, Watkins LR, et al. (2005) Medial prefrontal cortex determines how stressor controllability affects behavior and dorsal raphe nucleus. Nat Neurosci 8: 365-371.
69. Wiech K, Kalisch R, Weiskopf N, Pleger B, Stephan KE, et al. (2006) Anterolateral Pre-frontal Cortex Mediates the Analgesic Effect of Expected and Perceived Control over Pain. J Neurosci 26: 11501-11509.
70. Birbaumer N (2006) Breaking the silence: Brain-computer interfaces (BCI) for communication and motor control. Psychophysiology 43: 517-532.
71. Steinglass J, Walsh BT (2006) Habit learning and anorexia nervosa: A cognitive neuroscience hypothesis. Int J Eat Disord.
72. Taylor SF, Martis B, Fitzgerald KD, Welsh RC, Abelson JL, et al. (2006) Medial Frontal Cortex Activity and Loss-Related Responses to Errors. J Neurosci 26: 4063-4070.
73. Magno E, Foxe JJ, Molholm S, Robertson IH, Garavan H (2006) The Anterior Cingulate and Error Avoidance. J Neurosci 26: 4769-4773.
74. Somerville LH, Heatherton TF, Kelley WM (2006) Anterior cingulate cortex responds differentially to expectancy violation and social rejection. 9: 1007-1008.
75. Miller LS, Lachman ME (1999) The sense of control and cognitive aging: Toward a model of mediational processes.. In: Blanchard-Fields F, Hess T, editors. Social cogni-tion. New York: Academic Press. pp. 17–41.
76. Bardo MT, Donohew RL, Harrington NG (1996) Psychobiology of novelty seeking and drug seeking behavior. Behavioural Brain Research 77: 23-43.
77. Schultz W (2005) Behavioral Theories and the Neurophysiology of Reward. Annu Rev Psychol.
78. Schultz W (2001) Reward signaling by dopamine neurons. Neuroscientist 7: 293-302.
79. Schultz W (2002) Getting formal with dopamine and reward. Neuron 36: 241-263.
80. Schultz W, Dickinson A (2000) Neuronal coding of prediction errors. Annu Rev Neurosci 23: 473-500.
81. Mueller SC, Jackson GM, Dhalla R, Datsopoulos S, Hollis CP (2006) Enhanced Cognitive Control in Young People with Tourette’s Syndrome. Current Biology 16: 570-573.
82. Brown P, Marsden CD (1999) Bradykinesia and impairment of EEG desynchronization in Parkinson’s disease. Movement Disorders 14: 423-429.
83. Pessiglione M, Seymour B, Flandin G, Dolan RJ, Frith CD (2006) Dopamine-dependent prediction errors underpin reward-seeking behaviour in humans. 442: 1042-1045.
84. Munro, McCaul, Wong, Oswald, Zhou, et al. (2006) Sex Differences in Striatal Dopamine Release in Healthy Adults. Biological Psychiatry 59: 966-974.
85. Dayan P, Balleine BW (2002) Reward, motivation, and reinforcement learning. Neuron 36: 285-298.
86. Wolf R, Heisenberg M (1991) Basic organization of operant behavior as revealed in Drosophila flight orientation. Journal of Comparative Physiology A, Sensory, Neural, and Behavioral Physiology 169: 699-705.
87. O’Doherty J, Dayan P, Schultz J, Deichmann R, Friston K, et al. (2004) Dissociable roles of ventral and dorsal striatum in instrumental conditioning. Science 304: 452-454.
88. Krechevsky I (1937) Brain mechanisms and variability II. Variability where no learning is involved. Journal of Comparative and Physiological Psychology 23: 139-160.
89. McCormick DA (1999) Spontaneous Activity: Signal or Noise? Science 285: 541-543.
90. Lum CS, Zhurov Y, Cropper EC, Weiss KR, Brezina V (2005) Variability of swallowing performance in intact, freely feeding Aplysia. J Neurophysiol 94: 2427-2446.
91. de Ruyter van Steveninck RR, Lewen GD, Strong SP, Koberle R, Bialek W (1997) Reproducibility and Variability in Neural Spike Trains. Science 275: 1805-1808.
92. Gilden DL, Thornton T, Mallon MW (1995) 1/f noise in human cognition. Science 267: 1837-1839.
93. Maye A, Hsieh C, Sugihara G, Brembs B (2006, in prep.) Order in spontaneous behavioral activity in Drosophila.
94. Reynolds JNJ, Hyland BI, Wickens JR (2001) A cellular mechanism of reward-related learning. Nature 413: 67-70.
95. Poulet JFA, Hedwig B (2006) The Cellular Basis of a Corollary Discharge. Science 311: 518-522.
96. Brembs B, Lorenzetti FD, Reyes FD, Baxter DA, Byrne JH (2002) Operant reward learning in Aplysia: neuronal correlates and mechanisms. Science 296: 1706-1709.
97. Hawkins RD, Clark GA, Kandel ER (2006) Operant Conditioning of Gill Withdrawal in Aplysia. J Neurosci 26: 2443-2448.
98. Baxter DA, Byrne JH (2006) Feeding behavior of Aplysia: A model system for comparing cellular mechanisms of classical and operant conditioning. Learning and Memory 13: 669-680.
99. Putz G, Bertolucci F, Raabe T, Zars T, Heisenberg M (2004) The S6KII (rsk) gene of Drosophila melanogaster differentially affects an operant and a classical learning task. J Neurosci 24: 9745-9751.
100. Lorenzetti FD, Mozzachiodi R, Baxter DA, Byrne JH (2006) Classical and operant conditioning differentially modify the intrinsic properties of an identified neuron. Nat Neurosci 9: 17-29.
101. Lorenzetti FD, Baxter DA, Byrne JH. Both PKA and PKC are necessary for plasticity in a single-cell analogue of operant conditioning; 2006; Atlanta, Ga. USA. pp. 669.668.
102. Greengard P (2001) The Neurobiology of Slow Synaptic Transmission. Science 294: 1024-1030.
103. Greengard P, Allen PB, Nairn AC (1999) Beyond the Dopamine Receptor: the DARPP-32/Protein Phosphatase-1 Cascade. Neuron 23: 435-447.
104. Svenningson P, Greengard P (2006) A master regulator in the brain. The Scientist 20: 40.
105. Redgrave P, Gurney K (2006) The short-latency dopamine signal: a role in discovering novel actions? Nature Reviews Neuroscience 7: 967-975.
106. Menzel R, Leboulle G, Eisenhardt D (2006) Small Brains, Bright Minds. Cell 124: 237-239.
107. Brembs B (2003) Operant conditioning in invertebrates. Current Opinion in Neurobiology 13: 710-717.
108. Axton M (2006) Animal research and the search for understanding. Nature Genetics 38: 497-498.
109. Kleinfeld D, Griesbeck O (2005) From Art to Engineering? The Rise of In Vivo Mammalian Electrophysiology via Genetically Targeted Labeling and Nonlinear Imaging. PLoS Biology 3: e355.

 of science")












Comments are closed.