![]() Until 1986, it was thought that so-called optomotor responses, i.e., the tendency of all animals and humans to follow moving visual stimuli with their eyes or their bodies, were a prerequisite for gaze or trajectory stabilization: whenever the scenery in your gaze moves, you adjust gaze or body orientation to compensate for the shift. The main concept was that these behaviors were responses to external stimuli, e.g., shift to the left elicits a counter movement to compensate for the shift. Without such simple input-output processes, it was thought, it would be impossible to maintain course or gaze. However, in 1986, Wolf and Heisenberg reported on experiments in Drosophila fruit flies with drastically reduced optic lobes, the primary visual processing centers of insects. These flies with the double mutation rol (reduced optic lobes) sol (small optic lobes, see figure 1) were incapable of performing optomotor responses to rotating stimuli, but nevertheless fixated a stripe in their frontal visual field if they were allowed to control the position of that stripe with turning maneuvers that were simultaneously recorded. In other words, the flies could not see in which direction the stripe was moving, but nevertheless managed to move the stripe in a way that fixated it preferentially in front of the flies. Further experiments showed that the only way the flies could possibly accomplish this feat is by ‘trying out’ which turning maneuver led to the stripe staying in the frontal part of their visual field – an operant (or, as one would also say today, goal-directed) behavior. For more details on this and two related classic experiments from the Heisenberg lab see “The Importance of Being Active“.
Until 1986, it was thought that so-called optomotor responses, i.e., the tendency of all animals and humans to follow moving visual stimuli with their eyes or their bodies, were a prerequisite for gaze or trajectory stabilization: whenever the scenery in your gaze moves, you adjust gaze or body orientation to compensate for the shift. The main concept was that these behaviors were responses to external stimuli, e.g., shift to the left elicits a counter movement to compensate for the shift. Without such simple input-output processes, it was thought, it would be impossible to maintain course or gaze. However, in 1986, Wolf and Heisenberg reported on experiments in Drosophila fruit flies with drastically reduced optic lobes, the primary visual processing centers of insects. These flies with the double mutation rol (reduced optic lobes) sol (small optic lobes, see figure 1) were incapable of performing optomotor responses to rotating stimuli, but nevertheless fixated a stripe in their frontal visual field if they were allowed to control the position of that stripe with turning maneuvers that were simultaneously recorded. In other words, the flies could not see in which direction the stripe was moving, but nevertheless managed to move the stripe in a way that fixated it preferentially in front of the flies. Further experiments showed that the only way the flies could possibly accomplish this feat is by ‘trying out’ which turning maneuver led to the stripe staying in the frontal part of their visual field – an operant (or, as one would also say today, goal-directed) behavior. For more details on this and two related classic experiments from the Heisenberg lab see “The Importance of Being Active“.

Fig. 1: Side by side comparison of the brains of wild type (WT) and rol sol double mutant flies (click to enlarge).
Now Bahl et al. in the lab of Alexander Borst (a graduate student of Heisenberg‘s from 1982-84) have not only replicated this seminal study, they have done so in an impressively sophisticated and meticulous manner, avoiding many if not all of the methodical problems of the original experiments and adding exciting new findings to our body of knowledge along the way. Their fabulous publication constitutes a shining example of the power of combining today’s genetic technology with state-of-the-art behavioral physiology and computational modeling. Technically, it doesn’t get any better than that in our field today. A major procedural concern with the 1986 Wolf & Heisenberg paper had always been that none of the potential side-effects of the drastic double mutation rol sol were known. Granted, the optic lobes were vanishingly tiny in these mutants (see figure 1) and almost any visual capability seemed like a surprise, but which other processes were affected and in what way was completely unknown. Rather than almost ablating the entire optic lobes of the flies, Bahl et al.’s genetic manipulations specifically affected only two sets of interneurons, T4/5, in the flies’ visual system which the Borst lab had previously reported as abolishing all optomotor responses. Not only was it known that these highly specific manipulations abolished optomotor responses, it was also known why this happened: these neurons are a crucial component of the fly’s visual motion detector. Thus, these flies were exclusively motion-blind and all other faculties were unaltered. Amazingly, despite this highly specific manipulation, these flies behaved exactly like the rol sol flies: they did not respond to moving visual cues, but when allowed to control a black stripe with turning maneuvers, they kept the stripe in their frontal visual field. The authors not only replicated this aspect of the flies’ behavior, they also did so using walking flies, while the original study had examined flying flies, thus extending and generalizing the findings of Wolf and Heisenberg. Moreover, the authors found out, by careful and detailed dissection of the flies’ behavior, that optomotor responses improve fixation by increasing the tendency of the fly to move the stripe to the front of its visual field. For more details see the original publication or an excellent news-type article by Masson & Goffard. This news-article also emphasizes, btw, the relevance of fly work for our general understanding of gaze stabilization even in vertebrate systems.
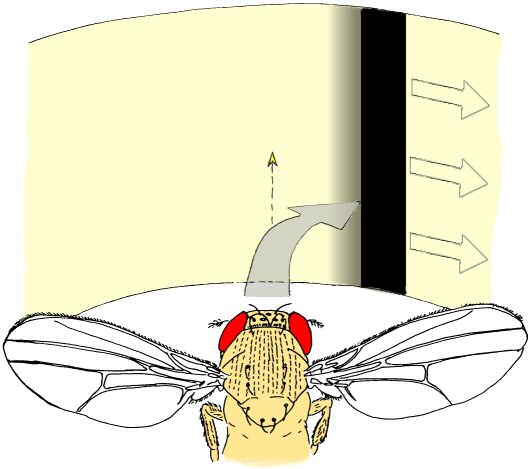
Fig. 2: “Inverting goggles” experiment. Whenever the fly attempts a turning maneuver, the fly’s visual panorama is rotated in the same direction. Fly drawing by R. Wolf.
Very much like Wolf & Heisenberg in 1986, Bahl et al. come to the conclusion that there is, in addition to the well-studied optomotor system, a less well-studied position system that acts in parallel during object fixation/course control. Specifically, and this is where they differ markedly from Wolf & Heisenberg, the authors assume that both systems operate according to more or less fixed input-output rules, i.e., “turn left on stimuli moving to the left” or “turn right if the bar is on the right”. However, the single most critical condition in the experiments by Wolf & Heisenberg was the one where a right turn of the fly led to a rotation of the visual field to the right, instead of to the left (the ‘inverting goggles‘ experiment, see figure 2). The rol sol flies showed no difference in stripe fixation between both conditions, whereas control flies took much longer to fixate the stripe under ‘inverting goggles’ conditions. This decisive experiment showed that “turn right when the stripe is on the right” (i.e., the conclusion Bahr et al. arrive at) cannot explain fixation in these motion blind flies. Had the rol sol flies performed this function exclusively, they would not have been able to fixate the stripe. The same then also holds for the flies from Bahr et al.: if their flies also fixate the stripe equally well in the regular as well as the inverted situation, their entire explanation for their results falls apart. Regrettably, Bahl et al. did not perform this very crucial, essential experiment, not in their flies, nor in the computer model they derived from their data. Hopefully, their next publication will contain these exciting experiments.
UPDATE: I have visited Axel Borst’s lab yesterday and they are working on the inverting goggles experiment. For now, the flies have trouble stopping the stripe from rotating at all, so there may be some technical problems using walking flies instead of flying flies for this kind of experiment, or simply the right parameters have not been found, yet. Either way, as I speculated, they are working on it!
Bahl, A., Ammer, G., Schilling, T., & Borst, A. (2013). Object tracking in motion-blind flies Nature Neuroscience, 16 (6), 730-738 DOI: 10.1038/nn.3386
Wolf, R., & Heisenberg, M. (1986). Visual orientation in motion-blind flies is an operant behaviour Nature, 323 (6084), 154-156 DOI: 10.1038/323154a0


 of science")












